Användbara noteringar om gastrering i groda
Användbara anteckningar om gastrering i groda!
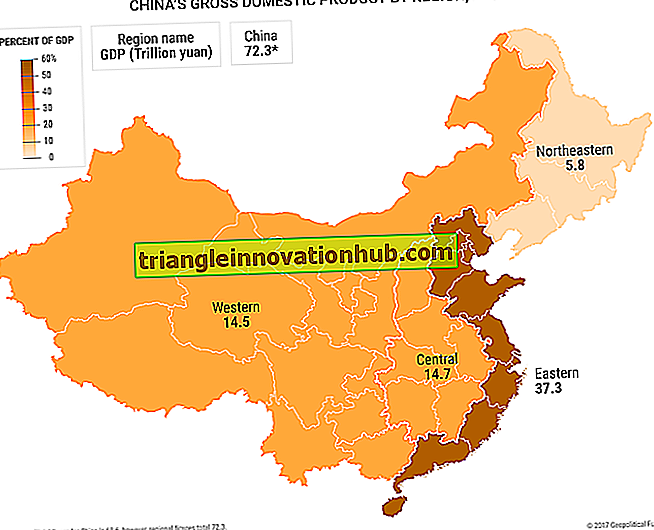
Gastrulering i groda:
Gastrering i processen med högintegrerad cell- och vävnadsmigration av potentiella endodermala och mesodermala områden till deras bestämda positioner i embryonets inre. Dessa rörelser är självbestämda och beroende av varandra och benämns morfogenetiska rörelser, skapar nya relationer och i slutändan ett triploblastiskt embryo. De cellulära beredningarna för dessa rörelser sker under klyvning.

Image Courtesy: mwilsonherps.files.wordpress.com/2011/11/ad-painted-frog.jpg
Amfibianfosteret genomgår en midblastula-övergång under vilken cellcykeln saktar ner (som en följd av cellens cyklens förvärv G1 och G2) blir celldelning en synkron, cellerna får förmågan att röra sig från sina ursprungliga positioner, och transkriptionen av nytt mRNA ses från kärnan för första gången i djurets liv. I Xenopus sker denna övergång omedelbart efter den tolfte klyvningen (Newport och Kirschner, 1982). Det förekommer tre typer av morfogenetiska rörelser vid amfibisk gastrering.
invagination:
I grodaembryon initieras gastrering vid den framtida dorsala sidan av embryot, strax under ekvatorn i gråmånens område. (Fig 11). Här sänker de marginala endodermala cellerna in i embryot och bildar sålunda en slitliknande blastopore. Dessa celler ändrar nu sin form och blir flaskformade.

Dessa kallas som flaskceller. Flaskcellerna håller kontakten med ytterytan med hjälp av cytoplasmatiska strängar medan huvudkroppen förskjuts mot embryonets insida. Därför börjar groddarna i marginalområdet nära ekvator av blastula. Här är endodermala celler inte så stora eller så ojämlika som den mest vegetabla blastomeren (fig 12 och 13).
Således, även om flaskcellerna kan vara ansvariga för att skapa det ursprungliga spåret, verkar den motiverande kraften här från de djupa lagren av marginalceller. Vidare verkar detta djupt lager av celler vara ansvarig för fortsatt migrering av cellerna i embryot.

Involution:
Nästa fas av gastrulation involverar invandringen av marginalzoncellerna, medan djurcellerna genomgår epibol och konvergerar vid blastopore. Vid att nå blastoporespetsen, svänger marginalcellerna inåt och reser längs inre ytan av de yttre cellerna (fig 11). Således förändras cellerna som utgör läppen av blastopore ständigt. De första cellerna som bildar dorsalipen är endodermala celler som invagineras för att bilda arkenteronets framkant.
Dessa celler blir senare faryngeala celler från förut. När dessa första celler passerar in i embryonets inre, blir blastoporeppen sammansatt av involverande celler som är föregångare till huvud mesodermen. Nästa celler som inverterar över blastopors dorsala läpp kallas chorda mesodermcellerna. Dessa celler kommer att bilda Notochord, ett övergående mesodermalt "bakben" som är nödvändigt för att initiera differentieringen av nervsystemet.
Epiboly:
När de nya cellerna går in i embryot, förskjuts blastokolen mot sidan mot den dorsala blastoporala läppen. Samtidigt förskjuts blastoporen vegetal och utvidgas när fler animaliska halvklotsceller konvergerar vid blastoporeppen. Den bredare blastoporen utvecklar laterala läppar och äntligen en ventral lip över vilken de ytterligare mesoderma och endodermala prekursorcellerna passerar. Med bildandet av ventralleppen har blastoporen bildat en ring runt de stora endodermala cellerna som förblir utsatta på ytan (fig 14).

Den återstående plåstret i endodermen kallas äggpluggen, och den också, i slutänden internaliserad (Fig 15). Vid denna tidpunkt har alla endodermala prekursorer förts in i embryonets inre, ectodermen har omringat ytan och mesodermen har förts mellan dem.